All published articles of this journal are available on ScienceDirect.

Purple Nonsulfur Bacteria for Greenhouse Gas Mitigation in Rice Systems: Mechanisms and Future Perspectives
Authors Info & Affiliations
Abstract
Introduction
Greenhouse gas (GHG) emissions from flooded rice soils, specifically methane (CH4) and nitrous oxide (N2O), are major contributors to agricultural global warming potential. While agronomic strategies, such as water and nutrient management, are used to mitigate, they often involve trade-offs among different gases. This review synthesizes effective GHG mitigation measures, including water management, rice straw management, chemical fertilizer management, rice cultivar selection, improved cultivation practices, and the role of purple nonsulfur bacteria (PNSB) as a multi-functional microbial solution for simultaneously mitigating CH4 and N2O emissions in rice systems.
Results
This review synthesizes current knowledge on GHG emissions from flooded rice systems and evaluates the emerging role of purple nonsulfur bacteria (PNSB) as a microbial strategy for mitigation. While conventional approaches such as water and fertilizer management can reduce emissions, their effectiveness is often constrained by agronomic trade-offs and environmental variability. Recent studies suggest that PNSB may contribute to reducing CH4 and nitrous oxide N2O emissions through multiple mechanisms, including modulation of carbon substrate availability, stimulation of methanotrophic activity, competition with methanogens, and enhancement of plant nutrient uptake.
Discussion
Compared with existing reviews, this study provides a mechanistic synthesis linking microbial metabolism with agronomic outcomes in rice systems. Evidence from laboratory and field studies indicates that PNSB inoculation can reduce CH4 emissions while improving plant growth. However, results remain inconsistent due to differences in soil type, management practices, and microbial strains. Key knowledge gaps include limited understanding of microbial interactions in situ, scalability of inoculation strategies, and long-term field performance.
Conclusion
Future research should integrate advanced analytical tools, including molecular techniques and data-driven approaches, to better resolve microbial processes and optimize PNSB-based biofertilizers. Overall, PNSB represents a promising but underexplored tool for sustainable rice production and climate change mitigation.
1. INTRODUCTION
Global surface temperatures during 2011–2020 were approximately 1.1 °C higher than during 1850–1900, due to increased greenhouse gas (GHG) emissions [1]. Agriculture, forestry, and land-use activities contribute approximately 22% of total global GHG emissions [2]. Rice cultivation alone emits more than 686 million tons of CO2-equivalent (CO2e), accounting for about 6.3% of total agricultural GHG emissions in 2021 [3]. Since 1750, atmospheric concentrations of CH4, N2O, and CO2 have increased by 47%, 156%, and 23%, respectively. Three gases are the primary GHGs emitted from rice fields. Over a 100-year timeframe, the global warming potentials of CH4 and N2O are approximately 28 and 273 times those of CO2, respectively [1, 4]. GHG emissions from rice fields are strongly influenced by water management, fertilizer application, rice straw management, and cultivar selection. Accordingly, various cultivation practices have been applied to mitigate GHG emissions [5–15]. Alternate wetting and drying (AWD) can reduce CH4 emissions but often leads to increased N2O emissions [16]. In rice straw management, returning rice straw to fields reduces CO2 emissions from open burning; however, incorporated straw serves as a substrate for methanogens, thereby increasing CH4 emissions [17, 18]. In addition, N2O emissions primarily result from nitrogen fertilizer application, with approximately 1–5% of applied N lost to the atmosphere as N2O [19, 20]. As shown, these approaches often involve trade-offs among emission reductions, crop yields, and soil fertility. As a result, there is growing interest in complementary biological strategies that can enhance both environmental and agronomic outcomes.
Soil microorganisms, particularly methanogens, play a crucial role in CH4 emissions from flooded rice fields. Organic matter-degrading bacteria produce acetate, CO2, and hydrogen (H2), which serve as substrates for methanogens, resulting in CH4 emissions [21]. Conversely, anaerobic methanotrophic archaea and methanotrophs oxidize CH4 under aerobic conditions, using it as a carbon source [22], and can reduce CH4 emissions by 60–80% in rice fields [23, 24]. Similarly, N2O emissions arise as by-products of microbial nitrification and denitrification. Under aerobic conditions, Nitrosomonas and Nitrobacter oxidize ammonium (NH4+) to nitrate (NO3-), whereas anaerobic bacteria reduce NO3- to dinitrogen (N2). During these processes, N2O is released as an intermediate prior to the completion of oxidation or reduction [25], contributing approximately 70% of global N2O emissions [26].
Beneficial microorganisms play essential roles in nutrient supply, plant growth promotion, and yield improvement in agricultural soils [27, 28]. Application of nitrogen-fixing bacteria as biofertilizers can reduce chemical nitrogen fertilizer use by up to 50% [29, 30], thereby mitigating N2O emissions. Purple nonsulfur bacteria (PNSB) exhibit strong potential for GHG mitigation through biological nitrogen fixation and nitrogen supply to rice plants [31]. PNSB strains such as Rhodopseudomonas palustris TN114 and PP803 have been shown to reduce CH4 emissions in rice fields [32, 33] and CO2 emissions [34]. Moreover, PNSB can grow under both favorable and stressful conditions, including saline, acid sulfate, and nutrient-poor soils [35–43]. Therefore, PNSB represents a promising approach for low-emission rice cultivation toward sustainable agriculture. However, the mechanisms and application strategies of PNSB for GHG mitigation in rice production remain insufficiently understood. Therefore, this review discusses current mitigation measures, mechanisms, and future prospects of applying PNSB to reduce GHG emissions in flooded rice systems. This review aims to (i) synthesize current knowledge on GHG emissions from rice systems, (ii) critically evaluate microbial mitigation strategies with a focus on PNSB, (iii) elucidate the underlying mechanisms of GHG reduction, and (iv) identify key research gaps and future directions for the application of PNSB in sustainable rice production.
2. CURRENT STATUS OF GHG EMISSIONS FROM FLOODED RICE SOILS
Rice cultivation contributes approximately 1.5% of total global GHG emissions and accounts for 48% of agricultural GHG emissions [44]. Among major crops, rice exhibits the highest CH4 emissions and ranks second in N2O emissions [3]. In 2022, global CH4 and N2O emissions from rice cultivation reached 24,384 kt and 183 kt, respectively, representing increases of 8.28% and 25.8% compared to 20 years earlier. Asia accounted for the largest share, emitting more than 21,397 kt CH4 (87.8%) and 146 kt N2O (88.7%) of total global emissions [3]. Average CH4 emissions from rice fields are approximately 1.98 kg ha-1 day-1, with the highest values observed in Vietnam (4.32 kg ha-1 day-1) and the lowest in India (0.65 kg ha-1 day-1). CH4 emissions from rice fields are closely associated with irrigation and organic matter management. Fields irrigated only by rainfall during the dry season or managed under soil moisture saturation emit 30–44% less CH4 compared with continuously flooded fields, while off-season straw incorporation can reduce emissions by up to 48% [45]. Average seasonal CH4 emissions are approximately 43.19 kg ha-1 season-1 [46], with annual emissions ranging from 272 to 614 kg ha-1 year-1 depending on region, while N2O emissions range from 1.19 to 1.29 kg ha-1 year-1 [17]. Rice cultivation contributes approximately 11% of agricultural N2O emissions worldwide [47], and these emissions are projected to increase by 35–60% by 2030 due to fertilizer management strategies and increased chemical nitrogen fertilizer use [48]. As can be seen, methane emissions from rice fields vary widely across studies, reflecting differences in soil properties, water management, organic inputs, and climatic conditions. For instance, higher emissions are commonly observed in continuously flooded systems with substantial organic matter inputs, as these conditions promote methanogenic activity. In contrast, practices such as intermittent drainage reduce CH4 production by introducing aerobic phases that suppress methanogens and stimulate methanotrophs. These variations highlight that emission rates are not solely site-specific but are strongly influenced by the interaction between management practices and microbial processes.
3. MITIGATION MEASURES FOR GHG EMISSIONS IN RICE FIELDS
Mitigation strategies for greenhouse gas (GHG) emissions in rice systems have primarily focused on water management, crop residue handling, fertilizer optimization, and cultivation practices. While these approaches are widely studied, their effectiveness varies considerably depending on environmental conditions, soil properties, and management interactions. Importantly, most strategies involve trade-offs between methane (CH4) and nitrous oxide (N2O) emissions, making it necessary to evaluate their overall impact in terms of Global Warming Potential (GWP) rather than individual gases.
3.1. Irrigation Management
Flooded rice fields create favorable anaerobic conditions for methanogenic activity. Mid-season Drainage (MD), intermittent irrigation, irrigation scheduling based on soil water potential, and saturated soil irrigation have been shown to reduce CH4 emissions by 30.0–84.7%, but can increase N2O emissions by up to 305.1% and reduce rice yield by 8.10–32.1% compared with continuous flooding [49, 50]. Although N2O contributes only about 5% to global warming potential (GWP), CH4 accounts for approximately 95%; therefore, MD can reduce GWP by 41–70% [51, 52].
Among various strategies, Alternate Wetting and Drying (AWD) is consistently identified as an effective method for reducing CH4 emissions. The primary mechanism involves periodic soil aeration, which limits the availability of substrates for methanogens and promotes methanotrophic activity [53–55]. In brief, GHG intensity and GWP decrease by 26.0–38.1% and 5.32–73.0%, respectively, while irrigation water use can be reduced by up to 70% without affecting rice yield [16, 54, 56–63], or with only a slight yield reduction of 1.56–11.6% [10, 64, 65]. In contrast, Arai [66] reported that AWD enhanced carbohydrate translocation from leaves to panicles, increasing rice yield by 9%. During drainage periods, surface soils become drier, and oxygen availability increases, creating unfavorable conditions for methanogens while stimulating organic matter oxidation and reducing dissolved soil organic carbon (SOC), thereby decreasing substrates for CH4 production and enhancing methanotrophic activity. However, alternating aerobic and anaerobic layers promotes both nitrification and denitrification, resulting in increased N2O emissions [16]. Combining AWD with deep fertilizer placement has been shown to reduce N2O emissions and GWP [67]. In a long-term monitoring over four consecutive seasons, Loaiza et al. [68] demonstrated that AWD not only saved water and reduced CH4 emissions but also reduced N2O emissions by 12–70% through improved soil moisture management during fertilizer application. As can be seen, the benefits of AWD are not uniform across studies. While CH4 reductions are generally substantial, N2O emissions often increase due to alternating nitrification–denitrification processes under fluctuating redox conditions.
In summary, irrigation control effectively reduces CH4 emissions during the dry season by suppressing methanogenic activity and enhancing methanotrophs, while conserving irrigation water [10, 69]. Proper soil moisture management, particularly by minimizing soil cracking during drainage, can also reduce N2O emissions by limiting the availability of nitrification and denitrification substrates [70]. However, further research is needed to clarify N2O emission dynamics under different irrigation regimes, as well as to address yield risks, rainy-season performance, and irrigation infrastructure investment for large-scale adoption [71].
3.2. Rice Straw Management
Rice straw residues are commonly burned in fields, incorporated into soil, used for bioenergy generation, mushroom cultivation, livestock feed, or composting [72]. Both open burning and straw incorporation into soil increase GHG emissions [73]. Returning straw to rice fields after harvest generally has little effect on N2O emissions but significantly increases CH4 emissions and GWP [6, 74]. Guo et al. [75] reported that straw incorporation increased CH4 and N2O emissions by 47.5% and 41.2%, respectively, and rice yield by 4.10%. GHG emissions associated with straw return depend on soil properties such as C/N ratio, available and total nitrogen, pH, straw incorporation methods, and cultivation practices. He et al. [6] suggested that returning straw at a depth of≥5 cm with rates >7.5 t ha-1 could reduce GWP. Overall, straw incorporation substantially increases CH4 emissions and can influence N2O emissions, thereby increasing GWP [6, 75]. The literature shows considerable variability in the effects of straw management, reflecting differences in incorporation timing, depth, and environmental conditions.
Removing straw from fields can reduce GWP by up to 45%, but negatively affects SOC, soil quality, and rice yield. Therefore, returning an appropriate amount of straw is considered an optimal strategy to reduce emissions while maintaining soil quality and yield [76]. Combining straw incorporation with AWD irrigation reduced CH4 emissions by 36–63% compared with straw incorporation under continuous flooding, with minimal effects on N2O emissions. Thus, straw incorporation should be combined with AWD to achieve effective GHG mitigation in rice cultivation [76, 77]. Surface straw mulching can also reduce CH4 emissions, particularly under water-managed conditions, due to reduced SOC availability [78]. Overall, managing fresh straw effectively contributes to GHG mitigation; however, contradictory findings remain, and challenges persist in persuading farmers to adopt alternative straw management practices [79]. Therefore, future research should focus on identifying context-specific straw management practices that balance GHG mitigation with soil sustainability.
3.3. Chemical Fertilizer Management
The amount of nitrogen (N) fertilizer applied to rice fields is directly associated with N2O emissions from nitrification and denitrification. Application of chemical N fertilizers increases N2O emissions by 72.5–311.1% due to the creation of favorable conditions for the activity of nitrifying archaea and bacteria [74], while simultaneously stimulating CH4 oxidation by methanotrophs, thereby reducing CH4 emissions by 7.20–49.0% and decreasing Global Warming Potential (GWP) by 13.5–17.9% [80, 81]. However, N fertilization also increases rice plant biomass and stimulates methanogenic activity by providing additional root exudates as carbon sources, thereby increasing CH4 emissions [20, 82, 83].
3.3.1. Microbial Mechanisms
The type of N fertilizer also influences GHG emissions. Substituting conventional urea with ammonium sulfate reduces CH4 emissions by stimulating the activity of sulfate-reducing bacteria, which compete with methanogens for hydrogen (H2), thereby suppressing CH4 production. However, this practice increases N2O emissions by enhancing denitrification rates [69]. Partial replacement of chemical fertilizers with biochar has been identified as an effective strategy for reducing GHG emissions in rice fields. Biochar application reduces cumulative N2O emissions by 15–32% over a 90-day period [84]. In addition, appropriate annual biochar application rates can reduce CH4 emissions by up to 89.6%, N2O emissions by 10.0–90.1%, and GWP by 21.5–35.7%, while increasing dissolved soil organic carbon (SOC), total soil nitrogen, and rice yield by 16.7–24.3% [7, 11, 85–88]. The primary mechanism underlying CH4 mitigation following biochar application is associated with increased soil pH and porosity, which reduce methanogen populations while enhancing the abundance and activity of methanotrophs [14, 89–91]. This effect is linked to biochar-mediated regulation of functional gene diversity, such as mcrA in methanogens and pmoA in methanotrophs. Biochar also regulates N2O emissions by suppressing key functional genes involved in the nitrogen cycle, including narG and nosZ, thereby reducing denitrification rates, nitrogen loss, and N2O emissions [92, 93]. However, biochar application increased N2O emissions by 30-190% due to creating a favorable environment for bacterial nitrification. In addition, the highly labile C/N ratio and the increase in soil pH associated with biochar application can enhance N2O emissions during denitrification [94, 95].
3.3.2. Agronomic Outcomes and Management Implications
From an agronomic perspective, optimizing nitrogen use efficiency is critical for reducing GHG emissions while maintaining crop productivity. Deep placement (subsurface application near rice roots) of urea in granular form reduces CH4 emissions by 9–49% and N2O emissions by 12–48.97% [96, 97], while increasing rice yield by 21% and nitrogen use efficiency by 58% [8, 54]. Similarly, deep placement of dissolved urea effectively reduced cumulative N2O and CH4 emissions by 15–28% and 43–67%, respectively, completely prevented ammonia volatilization under mid-season drainage conditions, and increased rice yield by 13–14% [15]. The use of slow-release urea or dissolved urea combined with deep placement reduced CH4 emissions by affecting CH4 production, oxidation, and transport processes. This practice stimulates rice root development in deeper soil layers, thereby increasing oxygen availability in the rhizosphere. As a result, CH4 oxidation is enhanced in deeper soil layers, and photosynthates are preferentially allocated to rice stems and leaves rather than roots. Consequently, methanogen populations are suppressed, whereas nitrogen inputs stimulate the growth and activity of methanotrophs, leading to reduced CH4 emissions but increased N2O emissions [96, 98]. Split application of urea according to crop demand, combined with deep placement, improves nitrogen use efficiency and reduces substrate availability for nitrification and denitrification processes. Consequently, N2O emissions from flooded rice fields are reduced during inundated growth stages [8, 97, 99].
Appropriate fertilizer management provides positive effects on GHG mitigation in rice fields. However, the interactions between nitrogen inputs and soil microbial communities require further investigation. Moreover, promoting the use of slow-release fertilizers or deep placement techniques among farmers remains a major practical constraint [79, 100].
3.4. Improved Cultivation Practices
Different cultivation practices, such as conventional tillage, shallow tillage, No-Tillage (NT), direct-seeded rice (DSR), or transplanted rice, significantly influence GHG emissions [79, 101]. NT reduces total GHG emissions by 45–51% [102] but increases N2O emissions by 65–300% [103] compared with deep tillage (20–22 cm) or shallow tillage. The use of DSR systems reduces CH4 emissions by 10.0–96.8% compared with transplanting [5, 104], without significantly affecting rice yield [105, 106]. In addition, large-scale farming systems are associated with 11% lower GHG emissions than smallholder farming [107].
Crop rotation also contributes to GHG mitigation [106]. CH4 and N2O emissions were reduced by 30.5% and 18.7%, respectively, under a rotation of one sesame crop followed by two rice crops [108]. Rotations involving peanut, soybean, or sweet potato with wheat–maize reduced N2O emissions by 30%, 42%, and 49%, respectively, due to lower nitrogen fertilizer application compared with monoculture controls [109]. The use of mulching films in crop rotation systems significantly reduced CH4 emissions by up to 59.2% [110] by decreasing functional microbial diversity and fermentative bacterial communities. Mineral mulching has also shown GHG mitigation effects comparable to those of agricultural plastic films (Table 1) [111].
| Cultivation Practice | CH4 Emissions | N2O Emissions | GWP | Rice Yield | References |
|---|---|---|---|---|---|
| NT | ↓ 57.3% | ↑ 69.7% | ↓ 29.46% | ↑ | Gangopadhyay et al. [112] |
| NT | ↓ 21.1–52.6% | ↑ 79–101% | ↓ 36.4% | ↓ 8.4% | Yang et al. [14] |
| NT | ↓ 18.3% | ↑ 28.9% | ↓ 27.7% | No difference | Guo et al. [75] |
| DSR | ↓ 47.0% | Not significant | ↓ 46.4% | No difference | Susilawati et al. [113] |
| DSR | ↓ 82.3–87.2% | ↑ 60–70% | ↓ 44.0–47.0% | ↓ 11.0% | Gupta et al. [114] |
| DSR + short-duration varieties | ↓ 46.1% | Not significant | ↓ 28.9–53.2% | No difference | Xu et al. [115] |
NT significantly reduces CH4 emissions by up to 57.3% due to reduced soil disturbance and slower hydrolysis of organic matter, thereby lowering substrate availability for methanogenic activity. However, N2O emissions increase for two main reasons: (i) soil stratification causes nitrogen fertilizers to accumulate primarily in the surface soil–water layer, providing substrates for nitrification and denitrification processes, thereby stimulating N2O emissions; and (ii) increased soil compaction reduces water-holding capacity, leading to faster soil drying and higher N2O fluxes. Compacted soils also restrict oxygen diffusion, further enhancing denitrification processes [14]. DSR practices also effectively reduce CH4 emissions by shortening the flooding period, promoting aerobic conditions in rice fields, and lowering rice root biomass. Reduced root exudation decreases SOC availability, thereby suppressing methanogen activity while enhancing CH4 oxidation by methanotrophs, thereby reducing CH4 emissions [104]. However, N2O emissions increase only slightly due to the counterbalancing effects of two processes: (i) aerobic conditions stimulate nitrification; and (ii) peak N2O emissions during rapid rice growth deplete soil nitrogen, improving nitrogen use efficiency in subsequent fertilizer applications and thereby limiting further N2O emissions [106, 113].
In summary, improved cultivation practices reduce CH4 emissions and overall GHG emissions in rice production but may increase N2O emissions. DSR systems still face several challenges, including shallow root systems that weaken soil anchorage and increase lodging risk, enhanced weed competition, increased disease pressure, and potential yield reductions [72, 116, 117]. With respect to crop residue accumulation and soil organic matter dynamics, more accurate assessments are required to evaluate the mitigation effectiveness of NT practices, proper implementation methods, and yield losses associated with soil-borne pathogens [118]. In addition, local climatic and soil conditions must be carefully considered when developing effective cultivation management strategies [119].
3.5. Rice Varieties
Rice varieties exhibit varying levels of GHG emissions. The variety BRRI Dhan29 produces relatively high emissions, whereas Binadhan-17 shows lower emissions, with corresponding CH4 emissions of 112.67 and 81.37 kg ha-1 season-1, respectively [57]. Low-emission rice varieties generally produce lower concentrations of dissolved SOC, NH4+, and NO3- in the soil [120]. Accordingly, high-yielding drought-tolerant varieties (7Y88, 7Y370) and salinity-tolerant varieties (Inpari 32, Ciherang, and Biosalin 1) emit less GHG than conventional varieties due to higher oxygen transport capacity, which creates more aerobic soil conditions. In addition, lower dissolved SOC provides less substrate for methanogens, thereby reducing CH4 emissions [64, 121]. Rice varieties with larger root biomass tend to emit less CH4 because more extensive root systems increase oxygen diffusion into the soil, thereby enhancing CH4 oxidation [122, 123]. Hybrid rice varieties reduce cumulative CH4 emissions by 18.6% due to greater root biomass, which results in a 93.4% increase in methanotroph abundance in soil compared with inbred varieties [124]. Lower shoot biomass and reduced stem–leaf rigidity are also associated with lower CH4 emissions [125]. Baruah et al. [126] further reported positive correlations between leaf area, leaf number, tiller number, root dry weight, soil SOC content, and the potential for CH4 and N2O emissions from rice plants. Improved, high-yielding, short-duration rice varieties such as OM18, OM2517, OM5451, and GKG 9 reduce CH4 emissions by 19–44% without affecting N2O emissions under flooded cultivation conditions, primarily due to genotypic differences [12].
The use of short-duration rice varieties (95–115 days) reduces CH4 emissions [127] because earlier maturity shortens the flooding period and reduces CH4 transport through rice tissues. However, N2O emissions increase due to the stimulatory effects of root exudates on soil nitrification processes [57]. In addition, early and prolific tillering can increase CH4 emissions during vegetative growth, as most CH4 is emitted through rice leaves and aerenchyma tissues [63, 106]. Long-duration rice varieties also show variable emission levels depending on stem, leaf, and root biomass. In India, the rice varieties IR64 and Number 9 exhibited lower GHG emissions than other varieties, with reductions of 21% and 34% in CH4 and N2O, respectively [128].
In summary, rice varieties differ in their GHG emission potential depending on growth duration, root–shoot biomass, and soil SOC levels. Short-duration, high-yielding, and hybrid rice varieties generally exhibit lower GHG emission potential than long-duration and traditional varieties. However, further research is required to elucidate the relationships among growth duration, genetic traits, environmental conditions, water management, and GHG emissions in rice varieties [12, 129].
Across all mitigation strategies, a consistent pattern emerges: individual practices rarely provide universal solutions due to complex interactions among soil, plants, and microbes. Most approaches reduce CH4 emissions but may increase N2O emissions, emphasizing the importance of evaluating mitigation strategies based on net GWP rather than single gases.
These limitations highlight the need for integrative approaches that combine agronomic management with microbial strategies, such as the use of purple nonsulfur bacteria. By targeting underlying biogeochemical processes, microbial interventions may help overcome trade-offs observed in conventional practices and provide more balanced, sustainable GHG mitigation in rice systems.
4. MICROBIAL APPROACHES FOR MITIGATING GREENHOUSE GAS EMISSIONS FROM FLOODED RICE SOILS
Methanotrophs are capable of oxidizing up to 80% of the CH4 produced in rice fields [24]. Accordingly, the application of methanotrophic bacteria such as Methylosinus sp. and Methylocystis sp. reduced CH4 emissions by 17–44%, which was associated with increased copy numbers of the pmoA gene, without affecting N2O emissions or rice yield in pot experiments using biogas effluent–based biofertilizers [13]. In addition, the application of Methylococcus capsulatus to soil, roots, and via foliar spraying reduced CH4 emissions by 31–60% and N2O emissions by 34–50%, while increasing grain yield by up to 39% [130]. Furthermore, the application of Aspergillus strains AMF1 and AMF2 and Trichoderma 1614 in rice fields with incorporated straw reduced soil SOC content and CH4 emissions [131]. The cable bacterium Electronema sp. GS increased soil sulfate concentrations fivefold, thereby stimulating sulfate-reducing bacteria that compete with methanogens for substrates, resulting in a 93% reduction in CH4 emissions compared with the control [132].
Most microbial applications reduce one or two GHG types. In contrast, PNSB has been shown to reduce multiple GHGs simultaneously in rice fields. In simulated microaerobic light conditions during rice straw decomposition, all 45 PNSB strains tested reduced CH4 emissions in saline soils [33]. Among these, Rhodopseudomonas (R.) palustris reduced CH4 and CO2 emissions by 86.49–100% and 38–47%, respectively [34], and by up to 80% and 33% in heavy metal–contaminated soils [9]. During growth, PNSB increased soil pH to 7.80, exceeding the optimal range for methanogens, thereby reducing CH4 emissions. In addition, PNSB uses CO2 for photoheterotrophic growth, thereby reducing CO2 emissions. Application of biofertilizers derived from R. palustris reduced CH4 emissions by 24–28%, with maximum fluxes of 50–70 mg m-2 h-1 compared with controls, and increased rice yield by up to 48% in saline and organic soils. The dominance of PNSB competitively inhibited methanogens by competing for acetate and other nutrients, thereby reducing CH4 emissions [32]. R. palustris significantly reduced average CH4 emissions to 37.26 g m-2 season-1 compared with 49.48 g m-2 season-1 in control treatments, corresponding to a GHG reduction equivalent to 3.05 t CO2-eq ha-1 season-1. Moreover, PNSB enhanced carbon sequestration processes, increasing soil carbon content by 257%, equivalent to 17.45 t C ha-1 [28].
Although chemical fertilizers are necessary to improve rice yield in intensive cropping systems [133], only approximately 50% of applied nitrogen is absorbed by crops during the growing season, while the remaining nitrogen is lost as NO3-, N2O, N2, or NH4+ [134]. The N2O emission factor of nitrogen sources is approximately 0.57%, with urea reaching 0.72% [135]. Consequently, for every 100 kg of urea applied to rice fields, up to 0.72 kg of nitrogen is released as N2O. Therefore, reducing chemical nitrogen fertilizer application is an effective strategy for mitigating N2O emissions [136]. PNSB possess one or more nitrogenase enzymes that enable biological N2 fixation and the supply of NH4+ to crops under aerobic, anaerobic, or microaerobic conditions, in both light and dark environments. Notably, R. palustris TN110 harbors all three types of nitrogenase genes, molybdenum–iron (Mo–Fe), vanadium–iron (V–Fe), and iron–iron (Fe–Fe), and exhibited the highest N2 fixation capacity among 235 evaluated strains [9]. Rhodobacter (Rba.) sphaeroides strains S01 and S06, isolated from saline soils, also fixed N2 under microaerobic light and aerobic dark conditions, with fixation rates of 16.9 and 32.1 mg L-1, respectively [35]. In rice fields, Rhodopseudomonas spp. are the most prevalent PNSB, increasing NH4+ concentrations by 19.4% and enhancing nitrogen uptake by up to 64 kg ha-1 [31, 137, 138]. Therefore, PNSB are considered effective biofertilizers that can partially replace chemical nitrogen fertilizers [139], thereby reducing N2O emissions from rice fields.
PNSB mitigate CO2 and CH4 emissions through carbon sequestration processes, suppress methanogen development in soil [28], and reduce N2O emissions by enhancing plant nitrogen uptake and minimizing nitrogen losses from soil [136, 138]. In addition, PNSB possess phosphate- and potassium-solubilizing capabilities and produce plant growth–promoting substances such as indole-3-acetic acid, 5-aminolevulinic acid, and siderophores, thereby enhancing rice growth and yield under adverse conditions [31, 36–39, 42, 140].
As shown in Table 2, compared with other microbial groups, PNSB exhibit a unique combination of metabolic flexibility and ecological adaptability, allowing them to function under both aerobic and anaerobic conditions. This positions them as potential integrators of multiple biogeochemical processes rather than single-function agents.
| Microbial Group | Mechanism | Strength | Limitation |
|---|---|---|---|
| Methanotrophs | CH4 oxidation | Direct CH4 reduction | Oxygen dependent |
| Fungi | Organic matter decomposition | Carbon regulation | Indirect effect |
| PNSB | Multi-pathway regulation | Versatile, adaptable | Field validation limited |
Overall, current evidence indicates that PNSB function as integrators of carbon and nitrogen cycling rather than as single-target mitigation agents. Their ability to simultaneously influence multiple GHG pathways represents a key advantage over conventional microbial approaches. Nevertheless, further research is required to validate these effects across diverse field conditions and to better understand interactions with native microbial communities and agronomic practices.
5. MECHANISMS OF GREENHOUSE GAS MITIGATION BY MICROORGANISMS
Microbial mitigation of GHG emissions in flooded rice soils is primarily governed by changes in carbon and nitrogen cycling. These mechanisms involve a network of microbial interactions that regulate methane (CH4) production and oxidation, as well as nitrous oxide (N2O) generation and reduction under fluctuating redox conditions. In general, CH4 emissions are controlled by the balance between methanogenic archaea, which produce CH4 from organic substrates, and methanotrophic microorganisms, which oxidize CH4 under aerobic or microaerobic conditions. Similarly, N2O emissions arise from intermediate steps in nitrification and denitrification pathways, which are mediated by diverse microbial communities. In addition to these well-established processes, emerging evidence highlights the role of functional groups such as cable bacteria and purple nonsulfur bacteria (PNSB), which influence GHG dynamics through alternative pathways, including sulfur cycling, carbon assimilation, and nitrogen fixation. The following sections summarize these key microbial mechanisms and their contributions to GHG mitigation in rice systems.
Cable bacteria perform electrogenic sulfur oxidation (e-SOx), which increases sulfate reserves in soil and reduces CH4 emissions through two main pathways: (i) Competition with methanogens: Elevated sulfate concentrations stimulate the activity of sulfate-reducing bacteria, which compete with methanogens for acetate as a substrate; (ii) Synergistic interaction with methanotrophs: Sulfate acts as an electron acceptor in anaerobic CH4 oxidation, thereby enabling cable bacteria to activate CH4 consumption via accumulated sulfate products (Fig. 1) [141].
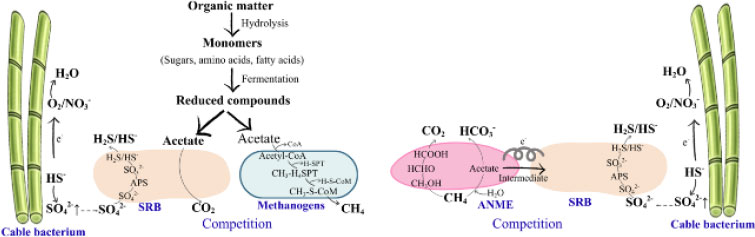
Methanotrophic CH4 oxidation occurs under a wide range of oxygen conditions. Under oxygen-rich conditions, aerobic methanotrophs assimilate approximately 50–60% of CH4-derived carbon into cellular biomass, while the remaining carbon is oxidized to CO2 and H2O through dissimilatory CH4 oxidation using O2 as the terminal electron acceptor via the enzyme methane monooxygenase (MMO) [142]. Under oxygen-limited conditions, anaerobic CH4 oxidation by methanotrophs is coupled with the reduction of NO3-, NO2-, iron, manganese, and sulfate [143]. Nitrate- and nitrite-dependent CH4 oxidation occurs intracellularly in methanotrophs through endogenous oxygen production during NO3-/NO2- reduction or by using NO3- as an alternative electron acceptor instead of O2 (Fig. 2) [144].
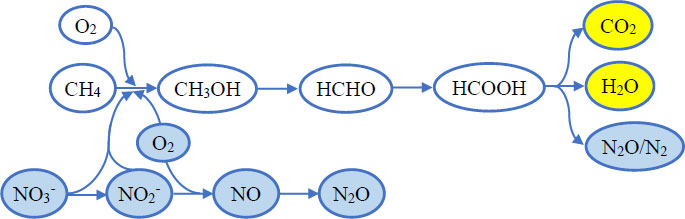
 Aerobic oxidation
Aerobic oxidation Anaerobic oxidation
Anaerobic oxidation Shared products of both processes.
Shared products of both processes.
The CH4 mitigation potential of PNSB was demonstrated as early as the study by Wertlieb and Vishniac [145], in which Rhodopseudomonas gelatinosa was shown to assimilate carbon from CH4 into cellular components and oxidize CH4 to CO2; however, at that time, the underlying mechanism was not clearly elucidated. During growth, PNSB utilizes a wide range of organic substrates under both aerobic and anaerobic conditions, including lignin, the main component of rice straw. While anaerobic bacteria degrade lignin and aromatic compounds to produce CH4 and CO2, R. palustris produces only CO2 during the degradation of benzene ring–containing aromatic compounds by using them as carbon sources via the enzyme benzoyl-CoA reductase (Fig. 3) [146]. After that, R. palustris PP803 was confirmed with methane reduction [32]. More recently, Wang et al. [28] also demonstrated that a R. palustris strain reduced CO2 and CH4. Nonetheless, there are currently no mechanistic studies involving GHG mitigation by PNSB. This reveals a remarkable scientific gap.
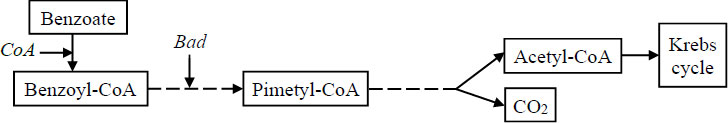
Anaerobic degradation pathway of aromatic benzoate compounds by R. palustris.
Source: Adapted from Hunter et al. [146].
Under photoheterotrophic conditions, PNSB compete with methanogens for acetate as a substrate [32], thereby reducing CH4 emissions. During this process, R. palustris converts approximately 22% of acetate into CO2, of which 68% is subsequently fixed into cellular biomass via the Calvin cycle [147]. Consequently, PNSB transform organic substrates into microbial biomass or generate GHG products with much lower global warming potential than CH4.
With respect to N2O mitigation, PNSB indirectly reduce emissions by enhancing nitrogen supply to rice plants through two primary mechanisms: (i) Mineralization of dead PNSB biomass, whereby nitrogen is gradually released for plant uptake following microbial decomposition; for example, Rhodobacter sp. biomass contains approximately 8.5% nitrogen on a dry weight basis [148]; (ii) Biological nitrogen fixation by living PNSB, which convert atmospheric N2 into NH4+ and enhance nitrogen uptake by rice plants [148]. In addition, PNSB reduces N2O emissions by utilizing NO3- as a nitrogen source through assimilatory nitrate reduction. Representative taxa involved in this process include Rhodobacter capsulatus, Rhodobacter sphaeroides, and members of the order Rhodobacterales. When NO3- is the sole nitrogen source, assimilatory nitrate reductase (Nas) is activated together with assimilatory nitrite reductase (Anir) to reduce NO3- in the cytoplasm, providing nitrogen for PNSB growth. R. capsulatus is capable of reducing hydroxylamine (NH2OH), a by-product responsible for N2O emissions during nitrification, into NH4+via Fe–S–containing enzymes (Fig. 4) [146]. Therefore, PNSB consume substrates and intermediate by-products of nitrification and denitrification processes to form microbial biomass, thereby minimizing N2O emissions in rice fields.
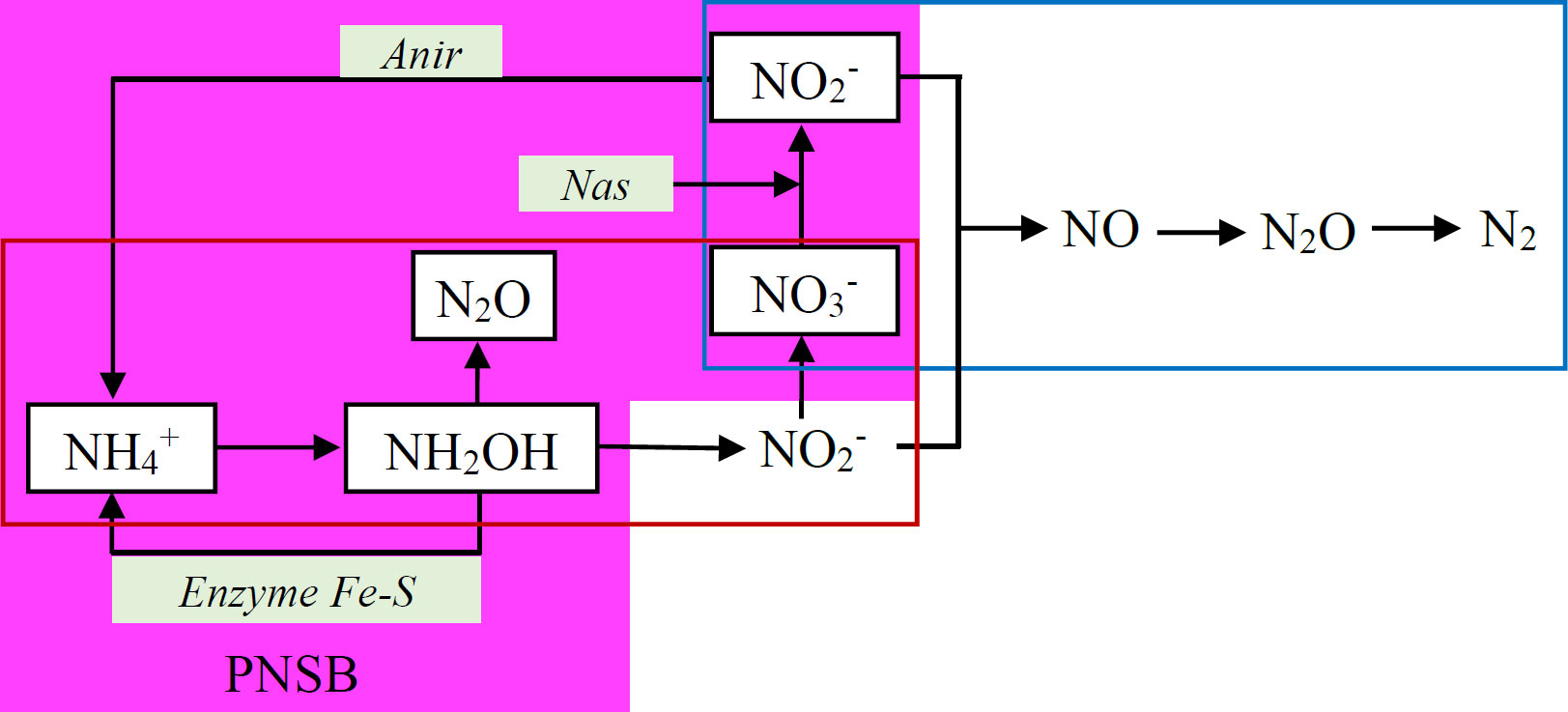
Mechanism of N2O mitigation by PNSB through assimilatory nitrate reduction.
Nas - assimilatory nitrate reductase; Anir - assimilatory nitrite reductase
 Denitrification;
Denitrification;  Nitrification;
Nitrification;  Assimilatory nitrate reduction
Assimilatory nitrate reduction
Source: Adapted from Hunter et al. [146].te reductase; Anir - assimilatory nitrite reductase
Denitrification; Nitrification; Assimilatory nitrate reduction
Source: Adapted from Hunter et al. [146].
6. CHALLENGES AND FUTURE DIRECTIONS FOR THE USE OF PNSB IN GREENHOUSE GAS MITIGATION
Current rice cultivation practices reduce certain GHGs while simultaneously increasing others, and trade-offs between CH4 and N2O emissions often determine the overall effectiveness in reducing global warming potential (GWP). In addition to management practices and policy factors, the simultaneous mitigation of CH4 and N2O remains a major challenge for developing effective GHG reduction strategies for rice fields in both the present and the future [5, 147]. The use of PNSB as biofertilizers has demonstrated strong potential to simultaneously reduce CH4 and N2O emissions [28, 136]. However, several practical constraints remain:
(i) Biomass production: PNSB requires specific nutrient media and optimal conditions such as light intensity, temperature, pH, and sterile environments for growth [148], making large-scale production challenging.
(ii) Storage and application methods: Liquid PNSB formulations are easy to culture but have short shelf lives and are difficult to transport. Solid formulations have longer storage stability but require suitable carrier materials, proper dilution according to recommended rates, and are highly susceptible to biotic and abiotic soil factors. Foliar application of PNSB is mainly effective for biological control and disease prevention, but does not fully exploit the multifunctional potential of PNSB in rice systems [42, 149]
(iii) Field-level adoption: Although the GHG mitigation potential of PNSB has been demonstrated experimentally, their application in low-emission rice production systems remains limited, and field performance is strongly influenced by environmental conditions [150]. In addition, the concepts of PNSB and GHG mitigation are still relatively new to farmers, limiting practical adoption. Economic assessments, including cost–benefit analyses of PNSB-based mitigation strategies, are therefore required [71].
(iv) Adoption and farmer awareness: Despite the promising potential of PNSB for GHG mitigation, their adoption at the farm level remains limited, partly due to low awareness and familiarity among farmers. PNSB-based technologies are relatively new compared with conventional inputs such as chemical fertilizers, and their benefits are not always immediately visible, which can reduce farmer confidence and willingness to adopt them.
To enhance the adoption of PNSB for GHG mitigation, cultivation of PNSB using rice straw fermentation water [151] should be further investigated and scaled up in field conditions to promote the development of indigenous PNSB biomass. This approach can reduce environmental impacts associated with external inoculum application and lower costs related to PNSB procurement and transportation. For example, application of Luteovulum sphaeroides increased soil NH4+ concentrations by 2.3 mg kg-1 in saline acid sulfate rice soils [140]. The use of indigenous Rhodopseudomonas spp. fixed 12.9–19.4 mg L-1 of nitrogen in acid sulfate rice soils, enabling a 25% reduction in chemical nitrogen fertilizer application [31]. Furthermore, field-scale experiments evaluating the simultaneous mitigation of CH4 and N2O emissions should consider multiple interacting factors, including rice straw management, cultivation practices, and fertilizer regimes, particularly in acid sulfate soils that impose multiple toxic constraints on microbial growth. Because PNSB thrives under anaerobic conditions and rice fields are predominantly anaerobic environments, the potential application of PNSB in flooded rice systems for effective GHG mitigation is highly promising. In addition, farmer training programs, participatory workshops, and on-farm demonstration trials can play a critical role in translating scientific knowledge into practical applications. In particular, demonstration plots comparing PNSB-treated and conventional practices under local conditions can provide tangible evidence of yield performance, reduced inputs, and environmental benefits. Such approaches have been shown to be effective in promoting the adoption of other biofertilizers and climate-smart agricultural practices.
Furthermore, advances in digital agriculture and data-driven technologies are increasingly enabling more precise monitoring and analysis of complex agroecosystems. These approaches offer new opportunities to better understand microbial dynamics and optimize sustainable management strategies [152].
CONCLUSION
Methane (CH4) emissions from rice fields are directly associated with the amount of rice straw and the duration of flooding, whereas nitrous oxide (N2O) emissions primarily depend on the rate, type, and application method of chemical nitrogen fertilizers. Therefore, straw management, irrigation control, nitrogen fertilizer management, and the adoption of improved cultivation practices are effective strategies for mitigating CH4 and N2O emissions from rice fields by influencing soil microbial communities. This review highlights the potential of purple nonsulfur bacteria as a multifunctional microbial tool for mitigating greenhouse gas emissions in rice systems. Unlike conventional strategies that target individual processes, PNSB influence multiple pathways simultaneously, including carbon substrate dynamics, methane production, and plant–microbe interactions. However, current evidence remains fragmented, and significant gaps persist in understanding field-scale performance, microbial interactions, and long-term impacts.
Future research should prioritize integrative approaches combining field experiments, molecular techniques, and data-driven modeling to better resolve the mechanisms and optimize application strategies. With further development, PNSB-based technologies could contribute to more sustainable and climate-resilient rice production systems.
AUTHORS' CONTRIBUTIONS
The authors confirm contribution to the paper as follows: P.N. and N.K.: Study conception and design; P.N.: Data collection; P.N., N.T., L.T., L.Q., T.N., M.B., N.P., L.X., H.K., D.X., and N.K.: Analysis and interpretation of results; P.N. and N.K.: Draft manuscript. All authors reviewed the results and approved the final version of the manuscript.
LIST OF ABBREVIATIONS
| GHG | = Greenhouse Gas |
| PNSB | = Purple Nonsulfur Bacteria |
| AWD | = Alternate Wetting and Drying |
| N2O | = Nitrous Oxide |
| SOC | = Soil Organic Carbon |
AVAILABILITY OF DATA AND MATERIALS
The authors confirm that the data supporting the findings of this study are available within the article.
FUNDING
This work was financially supported by Vietnam National University Ho Chi Minh City, Vietnam (Grant number B2024-16-02).
ACKNOWLEDGEMENTS
Declared none.

